



 Beeswax, Wax Glands
Beeswax, Wax Glands
Beeswax is a complex substance made up of wax esters, fatty acids and hydrocarbons.
By: Clarence Collision
Wax is used by honey bees to protect themselves against water loss through the integument and in the construction of combs. The major fractions of the cuticular wax were analyzed by gas-liquid chromatography and were shown to be qualitatively similar to those of comb wax (Blomquist et al. 1980). However, the composition of the cuticular wax of the honey bee is quantitatively different from that of the comb wax. The major component of the cuticular lipids is hydrocarbon, which comprises 58% of this wax. In contrast, hydrocarbon comprises only 13-17% of the comb wax, and monoester is the largest component (Tulloch 1971). Comb wax is produced by four pairs of glands within the abdomen while cuticular wax is likely produced by epidermal cells of the integument.
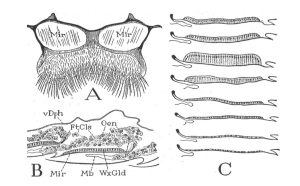
Figure 1.- The Wax Glands: A = Sternum of segment VI of worker, ventral, showing polished “mirrors” beneath wax glands, B = lengthwise section through two wax glands with overlying masses of fat cells and oenocytes. C = stages in the development and regression of a wax gland. Mir = mirror, WxGld = wax gland, FtCls = fat cells, Oen = oenocytes, vDph = ventral diaphragm, Mb = intersegmental membrane. (Snodgrass 1956).
Beeswax used in comb construction, comb repair, and capping of cells containing either pupae or honey, is secreted by worker bees on paired, smooth, oblong areas, called wax mirrors, located ventrally on abdominal segments four through seven (Figure 1A). On the dorsal side of each wax mirror is a layer of epithelial tissue called the wax gland (Sanford and Dietz 1976). Wax glands are merely specialized parts of the body-wall epidermis, which during the wax forming period in the life of the worker, become greatly thickened and take on a glandular structure (Figure 1C). The wax is discharged as a liquid through the mirrors and hardens to small flakes in the pockets between the mirrors and the long underlapping parts of the preceding sterna. After the wax-forming period the glands degenerate and become a flat layer of cells (Snodgrass and Erickson 1992).
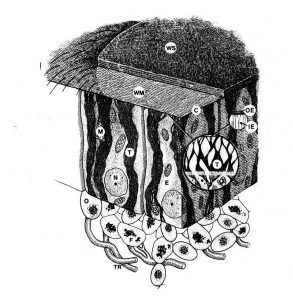
Figure 2. Diagram of the honey bee wax gland complex (ventral side up). WS = wax scale, WM = wax mirror, C = wax mirror cuticle, OE = outer epicuticle, IE = inner epicuticle, T = wax tubules, E = epidermal cell, N = nucleus, M = mitochondria, O = oenocytes, F = adipocytes and TR = tracheole. (Hepburn et al. 1991).
The wax gland complex of the honey bee worker (Figure 2) consists of three cell types, epithelial cells, oenocytes and adipocytes (fat body cells), which act synergistically to secrete wax, a complex mixture of hydrocarbons, fatty acids and proteins (lipophorins) (Cassier and Lensky 1995). Lying over each gland is a large cellular mass composed of fat cells and oenocytes (Figure 1B) (Snodgrass 1956).
Beeswax used in comb construction, comb repair, and capping of cells containing either pupae or honey, is secreted by worker bees on paired, smooth, oblong areas, called wax mirrors, located ventrally on abdominal segments four through seven.
In young bees that have just recently emerged from their cells, the cells of the wax gland are nearly square in shape, while in older bees, the cells are either elongated or completely degenerated (Figure 1C). There is a definite correlation between the age of the bee, the size (length) of the cells in the wax glands, and the gland’s ability to secrete wax (Turell 1974). In newly emerged bees the wax gland cells have large nuclei and do not have intercellular spaces (an open area between the cells). These cubical cells have an average height of 17 to 19 microns. The production of wax begins in workers that are slightly less than one week old. As secretory activity increases, the cells of the wax gland become tall and slender. Cells from a bee at peak wax production have a height of approximately 50 microns, and have developed large intercellular spaces. Wax glands are best developed and most productive in 12-18 day-old workers. After producing wax for a few days, the wax glands begin to degenerate and by the time the bee is ready to leave the hive to become a field bee, usually when it is about 21 days of age, the glands have completely degenerated. The cellular boundaries, which were distinct in the immature and active glands, have become indistinct, or even lacking, and the height of the gland has fallen to only three microns.
Beeswax is produced by metabolizing honey in fat cells associated with the wax glands and converting it to beeswax; workers cannot produce beeswax unless there are adequate honey stores in the colony. Workers also need to eat pollen during the first five to six days of their life in order to secrete wax later on, evidently because the protein in pollen is needed at that time for adequate fat cell development (Winston 1987). Wax is secreted primarily during warm weather when foraging is active. Workers actively engaged in secreting wax engorge themselves with honey and hang in festoons at or near the site of comb building. Drones and queens do not have abdominal wax glands.
To date, the mechanisms associated with wax synthesis and secretion are not fully understood. Piek (1964) showed that acetic acid is likely taken up by the oenocytes and that acetate is used for the synthesis of hydrocarbons. Blomquist et al. (1980) demonstrated that the incorporation of injected radio-labelled acetate into hexane extractable wax (cuticular wax) by worker honey bees not actively producing comb wax resulted in the recovery of much of the radioactivity in the hydrocarbon fraction. In bees actively producing comb wax, a higher percentage of radioactivity was recovered in the monoester fraction. A dramatic effect of age on the distribution of radioactivity from acetate into the various wax fractions from bees studied in the Summer months was observed. In bees from 11 to 18 days following emergence to adults, the major wax component synthesized was monoester, whereas in younger and older bees, hydrocarbon was the major wax component formed. Both in vivo and in vitro experiments using bees actively producing comb wax showed that the abdomen produced significant amounts of monoester, hydrocarbon, and other esters, whereas the thorax synthesized mostly hydrocarbon. These data show that the epidermal cells and wax glands each produce a wax with a distinct composition, and that the age and seasonal differences observed in wax synthesis are due to presence or absence of active wax glands.
There is a definite correlation between the age of the bee, the size (length) of the cells in the wax glands, and the gland’s ability to secrete wax.
The ultra-structure of the cells of the wax gland complex was studied in relation to the synthesis and secretion of beeswax (Hepburn et al. 1991). The hydrocarbon and fatty acid profiles of epidermal cells and oenocytes were determined in relation to the ages of the bees. Smooth endoplasmic reticulum (SER)** was absent from both epidermal cells and adipocytes (fat cells) in adult workers from the time they emerged until the end of wax secretion. The oenocytes were rich in SER. The hydrocarbon and fatty acid content of the oenocytes, averaged for bee age, closely matched that of newly secreted wax. The oencytes are the probable source of the hydrocarbon fraction of beeswax which is consistent with histochemical and autoradiographic data. The cyclical changes of organelles within the cells and chemical composition of the wax gland complex closely coincided with measured, age-related rates of wax secretion in the workers.
Hepburn et al. (1991) observed that SER is barely discernible in the oenocytes of newly emerged workers, but by the fourth day the volume density of these tightly packed tubules is high.
Similarly, there is a large increase in whole oenocyte volume and the relative volumes of the oenocytes and SER remain elevated throughout the secretory phase. By day 18, both oenocytes and SER begin to decrease with the simultaneous appearance of primary lysosomes (membrane enclosed organelles in the cell that contain an array of digestive enzymes) and autolytic vacuoles (autolytic- cell destruction through the action of its own enzymes and vacuoles are organelles containing debris at various stages of degradation). Lipid and protein droplets were never observed in the oenocytes and cellular organelles showed no evident cyclical changes associated with wax synthesis. The fat cells are characterized by an extensive plasma membrane reticular system, numerous mitochondria (organelles that function in energy production), peroxisomes (organelles that contain enzymes involved in metabolic reactions) and rough endoplasmic reticulum and a few small Golgi bodies. The Golgi bodies or Golgi complex take up and process secretory products from the endoplasmic reticulum and then either releases the finished product into the cell cytoplasm or secretes them outside of the cell. SER is notably absent in fat cells from adult emergence through foraging. Massive lipid droplets occupy approximately 60% of cell’s cytoplasm in the young worker but decrease substantially over the next few days. Like the oencytes, the fat cells also increase in volume prior to wax synthesis. During wax synthesis, glycogen stores are notably large and the plasma membrane reticular system is well developed. As synthesis wanes, lipid droplets increase in size while the other organelles either remain unchanged or show small decreases in size. The notable and dynamic feature of the oenocyte is the abundant SER whose rise and fall are synchronized with measured periods of secretion (Hepburn et al. 1991). The major role of the epidermis in the production of wax appears to be the development of an elaborate system of small transport tubules (Locke 1961).
The structure of the wax mirrors and the different types of cells were studied with scanning and transmission electron microscopy (Cassier and Lensky 1995). The outer face of the wax mirror shows a sub-regular, hexagonal pattern, each unit corresponding to an underlying epithelial cell. On perfectly clean wax mirrors, the covering epicuticle shows numerous holes or depressed areas from which the new biosynthesized wax masses exude, fuse and form irregular puddles. The sternal cuticle is particularly thin at the level of the mirror plates (two to four µm). It is composed of an outer trilaminate epicuticle, an homogeneous inner epicuticle and a two- or four-layered procuticle.
The main characteristic of the wax mirror is the presence of numerous pore canals containing microfilamentous structures or wax canal filaments (Locke 1961).
Beeswax when first secreted by the wax glands appears as a translucent white ellipsoidal flake. Production of beeswax in the honey bee is a process whereby many thin layers of wax are deposited on the wax mirror until a scale results. Dietz and Humphreys (1970) studied the structure of the wax scales with a scanning electron microscope. The customary shape of wax scales is due to the slightly recessed wax mirrors or plates which are situated below the wax glands. Essentially the wax scales produced by the wax glands of segments IV, V, and VI are somewhat similar in size and shape. Those originating with segment VII are not only smaller but also of different shape.
The mechanism by which beeswax penetrates the cuticle to form the layers of the scale has been subject of several investigations (Sanford and Dietz 1976). The extensive system of pore canals filled with filaments, believed to consist of wax extending up into the wax canals, is thought to form part of the transport mechanism which brings wax or its precursors near to the surface of the wax mirrors (Goodman 2003).
The cuticle of the wax mirrors has a stratified structure and the associated fibrous structures and pore canals certainly play a major function during the transit of the wax components from the wax glands to the exterior surface (Locke 1961). It has been hypothesized that the wax components are transported in a protein medium through the gland cells to the outer surface of the mirrors, where the molecules condense to form scales. Kurstjens et al. (1990) electrophoretically detected proteins in the wax scales and in the comb wax.
Cassier and Lensky (1995) with scanning and transmission electron microscopy showed the extrusion of wax droplets through the wax mirrors and for the first time large cisternae (fluid containing sac or cavity) of smooth endoplasmic reticulum in the epidermis. These cisternae are probably involved in the transit of wax from the oenocytes to the pore canal system. The cisternae can also convey apolipophorins from the hemolymph to the wax mirrors.
The elongate epithelial cells of the wax gland form a palisade layer. Intercellular spaces and infoldings of the plasma membrane delimit large spaces where twisted filamentous structures run and connect to those of the pore canals (Cassier and Lensky 1995). Beeswax is a composite mixture of hydrocarbons, esters, fatty acids (Hepburn et al. 1991) and proteins (Kurstjens et al. 1990). Because beeswax is hydrophobic it is probably transported from the wax glands to the mirrors by hemolymph lipophorins via the SER cisternae.
Based on biochemical and structural data, it is possible to suggest the contributions of each cell type to the wax gland complex. Oenocytes are involved in the secretion of the hydrocarbon fraction of the wax (Piek 1964; Blomquist and Ries 1979; Lambremont and Wykle 1979; Hepburn et al. 1991). The epithelial cell provided with ribosomes, polysomes, rough endoplasmic reticulum cisternae and electron-dense granules probably synthesizes a part of the protein fraction of the wax product, the other part coming directly from hemolymph through SER cisternae. Fat body cells provide plastic and energetic products. In contrast to previous workers, Cassier and Lensky (1995) found that the SER is well developed both in oenocytes and epidermal cells. In the epidermal cells, SER forms long cisternae parallel to the major axis, from the basal to the distal pole of the epithelial cells. They seem to be connected to extracellular spaces, thus forming a complete pathway to the wax mirror exterior.
Examination of the fine structure of the wax gland indicates that its function in beeswax secretion is either strictly transport or concentration of substances rather than producing them for release as in the case with many secretory cells (Sanford and Dietz 1976). Piek (1964) showed that the constituents of beeswax are synthesized in the fat cells and oenocytes, which are enzyme activated by an esterase, which could possibly catalyze wax production in the cuticle as well as in the epithelium. Piek concluded that esters are synthesized by fat cells and hydrocarbons and wax acids by the oenocytes. These wax precursors are then discharged into the wax gland (epithelium), concentrated and then pumped into the extracellular space.
The actual site of the final reactions that complete beeswax formation before it is secreted through the pore canals is not known. It may be assumed from its accumulation as a liquid layer on the wax mirrors, and from its solidification rapidly after secretion, that the final reactions that result in hardening of the wax take place after secretion.
Beewax is a complex substance made up of wax esters, fatty acids and hydrocarbons (Piek 1964; Tulloch 1970). Over 300 individual chemical components have been identified from pure beeswax (Tulloch 1980). Beeswax consists primarily of monoesters (35%), hydrocarbons (14%), diesters (14%), triesters (3%), hydroxymonoesters (4%), hydroxypolyesters (8%), free fatty acids (12%), acid esters (1%), acid polyesters (2%), free alcohol (1%) and unidentified (6%). It is this great diversity of composition that gives beeswax many unique properties (Goodman 2003) and keeps us from fully understanding the synthesis and secretion process.
References
Blomquist, G.J. and M.K. Ries 1979. The enzymatic synthesis of wax esters by a microsomal preparation from the honey bee Apis mellifera L. Insect Biochem. 9: 183-188.
Blomquist, G.J., A.J. Chu and S. Remaley 1980. Biosynthesis of wax in the honeybee, Apis mellifera L. Insect Biochem. 10: 313-321.
Cassier, P. and Y. Lensky 1995. Ultrastructure of the wax gland complex and secretion of beeswax in the worker honey bee Apis mellifera L. Apidologie 26: 17-26.
Dietz, A. and W.J. Humphreys 1970. Scanning electron microscopy study of the structure of honey bee wax scales. J. Georgia Entomol. Soc. 5: 1-6.
Goodman, L. 2003. Form and Function in the Honey Bee. International Bee Research Association, Cardiff, UK, 220 pp.
Hepburn, H.R., R.T.F. Bernard, B.C. Davidson, W.J. Muller, P. Lloyd, S.P. Kurstjens and S.L. Vincent 1991. Synthesis and secretion of beeswax in honey bees. Apidologie 22: 21-36.
Kurstjens, S.P., E. McClain and H.R. Hepburn 1990. The proteins of beeswax. Naturwissenschaften 77: 34-35.
Lambremont, E.M. and R.L. Wykle 1979. Wax synthesis by an enzyme system from the honey bee. Comp. Biochem. Physiol. 63B: 131-135.
Locke, M. 1961. Pore canals and related structures in insect cuticle. J. Biophys. Biochem. Cytol. 10: 589-618.
Piek, T. 1964. Synthesis of wax in the honeybee (Apis mellifera L.) J. Insect Physiol. 10: 563-572.
Sanford, M.T. and A. Dietz 1976. The fine structure of the wax gland of the honey bee (Apis mellifera L.). Apidologie 7: 197-207.
Snodgrass, R.E. 1956. Anatomy Of The Honey Bee, Comstock Publ. Assoc., Ithaca, NY, 2nd ed.
Snodgrass, R.E. and E.H. Erickson 1992. The anatomy of the honey bee. In The Hive And The Honey Bee, Dadant and Sons, Inc., Hamilton, IL. pp. 103-169.
Tulloch, A.P. 1970. The composition of beeswax and other waxes secreted by insects. Lipids 5: 247-258.
Tulloch, A.P. 1971. Beeswax: structure of esters and their component hydroxyl acids and diols. Chemy. Phys. Lipids. 6: 235-265.
Tulloch, A.P. 1980. Beeswax- composition and analysis. Bee Wld. 61: 47-62.
Turell, M.J. 1974. The wax glands of the honey bee. Am. Bee J. 114: 328-330.
Winston, M.L. 1987. The Biology Of The Honey Bee. Harvard University Press, Cambridge, MA, 281 pp.
Clarence Collison is an Emeritus Professor of Entomology and Department Head Emeritus of Entomology and Plant Pathology at Mississippi State University, Mississippi State, MS.
** Endoplasmic reticulum is a membrane system of folded sacs and interconnected channels within the cell’s cytoplasm that serve as a site for protein and lipid (fat) synthesis. Rough endoplasmic reticulum (RER) is in the form of flat bands covered with ribosomes that are responsible for the synthesis of many proteins. Smooth endoplasmic reticulum (SER) is tubular in form; serving as the site of lipid synthesis and carbohydrate metabolism.

